Часть 2. Начало статьи читайте в № 4, 2019 г.
Легочный микробиом и астма
Астма является наиболее распространенной респираторной хронической патологией в любой возрастной группе. Точные механизмы, которые приводят к различным клиническим фенотипам и эндотипам и присущим специфическим воспалительным процессам при астме, по-прежнему в значительной степени неизвестны. Несмотря на это, в свете современных публикаций можно думать о том, что барьерная функция слизистой оболочки носа и микробиота носа являются ключевыми элементами, связывающими респираторный дисбиоз с клеточными и молекулярными процессами, которые приводят к воспалению и поддерживают его. Повышенная проницаемость слизистой оболочки может, в свою очередь, способствовать перемещению бактерий в подслизистый слой, где происходит встреча антигена с соответствующими иммунными клетками и его распознавание, с последующим запуском локальных или системных реакций.
У здоровых людей микробиота легкого детерминируется в основном балансом иммиграции и элиминации с относительно небольшим вкладом от дифференциального коэффициента воспроизводства микробного сообщества [36].
Основными маршрутами микробной иммиграции в легкие являются микроаспирация, вдыхание воздуха и прямая дисперсия вдоль поверхностей слизистой оболочки. Из них микроаспирация, вероятно, является доминирующим маршрутом иммиграции.
Исследования микробиомов носа, полости рта, легкого и желудка у индивидуумов иллюстрируют микробиологическую непрерывность аэродигестивного тракта у здоровых взрослых, что обосновывает микробиологическую поддержку концепции о распространенности микроаспирации у здоровых людей [37, 38]. Таким образом, даже в норме постоянно наблюдается бактериальная контаминация нижних дыхательных путей. Очищение дыхательных путей происходит за счет мукоцилиарного клиренса, кашля. Известно, что при отсутствии инфекции здоровые люди кашляют примерно один раз в час, стимулируя продвижение бактерий в глотку, где они либо проглатываются, либо отхаркиваются.
Элиминация микроорганизмов (кроме кашля и мукоцилиарного клиренса) осуществляется разнообразным арсеналом врожденной и адаптивной иммунной защиты, который избирательно распознает некоторые микроорганизмы, вызывая их гибель или тормозя размножение. Доказано, что содержание иммунных клеток в легких связано с особенностями микробиоты [39].
В отличие от других систем дыхательные пути имеют гомогенную микробиоту, которая уменьшается в биомассе от верхних отделов к нижним. Здоровое легкое не имеет обособленного микробиома, но вместо этого содержит низкие уровни бактериальных сиквенсов, которые в значительной степени неотличимы от верхней дыхательной флоры [33].
Стремительное накопление информации о микробиоме дыхательных путей из-за выраженной гетерогенности исследований не позволяет сформулировать единую четкую концепцию роли микробиома при астме, поскольку микробиом может быть как фактором риска развития астмы, так защиты от нее. Патогенез астмы разнообразен и включает экспозицию аллергенов, другие экспосомы, факторы окружающей среды, взаимодействующие с генетикой во время беременности и в первые годы жизни. Микробный дисбиоз в дыхательных путях все чаще ассоциируется с частотой и тяжестью астмы. Более точному эндотипированию пациентов с астмой может помочь дальнейший анализ состава и метаболической активности индивидуального микробиома. При разных фенотипах астмы наблюдаются отличия в формировании бактериальной колонизации дыхательных путей определенным спектром бактерий, которые можно трактовать как возможности микробиома, определять время дебюта и тяжесть астмы, чувствительность к глюкокортикостероидам (ГКС) и макролидам, коморбидностям [40].
При нейтрофильном Th17-ответе наблюдается преобладание
Дыхательные пути пациентов с устойчивостью к ГКС и тяжелым течением заболевания колонизируются Moraxella сatarrhalis, Haemophilus или Streptococcus [42, 43].
У пациентов с ГКС-резистентной астмой в составе микробиома преобладали Neisseria, виды Haemophilus, виды Campylobacter и Leptotrichia, которые отсутствовали у пациентов с астмой, чувствительной к ГКС [44].
Использование ГКС, особенно сочетание ингаляционных и пероральных, связано с увеличением численности Proteobacteria и рода Pseudomonas и уменьшением численности видов Bacteroides, Fusobacteria и Prevotella [45].
Лечение ингаляционными глюкокортикостероидами было связано с изменениями профиля цитокинов и воспалительного ответа дыхательных путей на микробиоту. При атопической астме характер воспаления коррелировал с составом микробиоты и серьезностью обструкции дыхательных путей. У пациентов с тяжелой бронхиальной астмой наблюдались снижение концентрации ИЛ-2 и ИЛ-10, высокие концентрации ФНО-α и MIP-1β, колонизация Actinomyces, Neisseria menigiditis и Streptococcus pneumoniae.
У пациентов с более легкими вариантами бронхиальной астмы были значительно снижены концентрации ИФН-γ, ИЛ-17F и G-CSF, повышен ИЛ-2, отмечена колонизация видами Enterococcus. После терапии ингаляционными ГКС в течение 6 недель отмечалась положительная динамика в отношении бронхиальной обструкции, выявлено значительное увеличение видового разнообразия в фенотипах с выраженной и умеренной обструкцией — снижение численности патогенных видов, улучшение качественного состава микробиома [46].
Последние несколько лет привели к ключевым исследованиям, которые позволили четко связать изменения в микробиоме с повышенным риском развития астмы и аллергии. В рамках существующей концепции иммунной солидарности слизистых не только микробиом дыхательных путей несет ответственность за развитие воспаления, но и кишечный микробиом влияет на анатомически удаленные слизистые и иммунную систему в целом. Ассоциации между кишечной и респираторной микробиотой, называемые кишечно-легочной осью, играют центральную роль в динамических изменениях, происходящих при астме и аллергии [47].
«Здоровая» легочная микробиота характеризуется небольшой биомассой, но чрезвычайным разнообразием микробных видов. «Здоровая» кишечная микробита характеризуется одновременно и разнообразием микроорганизмов, и большой биомассой. Микробиота кишечника участвует в формировании лимфоидной архитектуры и иммунного ответа. Потеря кишечного микробного разнообразия (например, в результате вирусной инфекции или использования антибиотиков) может приводить к дисрегуляции ИЛ-17 или бактериальных механизмов элиминации, что потенциально может привести к дисфункции альвеолярных макрофагов и селекции микроорганизмов с патогенным потенциалом (обострение легочного заболевания).
Некоторые формы хронических заболеваний легких могут обостряться из-за транслокации и/или изменения бактериального содержимого кишечника. Воспалительное повреждение легочной ткани (например, вирусная инфекция или аспирация) могут утяжелить заболевание из-за воздействия на микробиоту легкого или кишечника. Вполне вероятно, что существует критическая взаимосвязь или равновесие между компонентами микробиома до тех пор, пока не возникнут возмущения, которые приводят к состоянию дисбиоза или «нездоровому» микробиому [48].
Микробиота кишечника питается и созревает в кишечном эпителии и участвует в иммунитете. Основная роль микробиоты легкого — ориентация и баланс аспектов иммунной и эпителиальной реакции [37].
Перенос акцента на роль эпителия нацеливает исследователей и клиницистов обращать большее внимание на защиту уязвимых дыхательных путей от воздействия окружающей среды и восстановление дефектных функций эпителия, а не только на подавление воспаления дыхательных путей или модуляции иммунного ответа [49].
Эпителиальное/мезенхимальное взаимодействие через экспрессию эпителиальных генов периостина (POSTN), регулятора хлористых каналов 1 (CLCA1) и ингибитора серпин-пептидазы B2 (SERPINB2) специфическим образом изменяется при астме и ассоциируется с ИЛ-13-связанным воспалением дыхательных путей [50]. ИЛ-13 продуцируется Т-хелперами (Th2), а также клетками врожденного иммунитета (Innate Lymphoid Cells, ILC2) [51].
Врожденные лимфоидные клетки
Врожденные лимфоидные клетки (Innate Lymphoid Cells, ILCs) — это группа лимфоцитов, которые вовлечены в быстрое цитокинзависимое реагирование организма во время воспалительного процесса [52]. В организме врожденные лимфоидные клетки (ВЛК) рассредоточены преимущественно в барьерных тканях, на границе внешней и внутренней среды (например, в слизистых). Больше всего ВЛК в месте максимальной концентрации всех иммунокомпетентных клеток нашего организма — в лимфоидной ткани слизистой оболочки пищеварительного тракта. Здесь ВЛК отвечают за контроль мирного сосуществования с населяющими слизистые желудочно-кишечного тракта бактериями «нормальной флоры». ВЛК в достаточном количестве обнаруживаются и в слизистых дыхательных путей [53].
На рис. 2 представлены три группы ВЛК в зависимости от их функций (адаптировано с сайта http://naukageek.blogspot.com/2014/06/innate-lymphoid-cells.html).
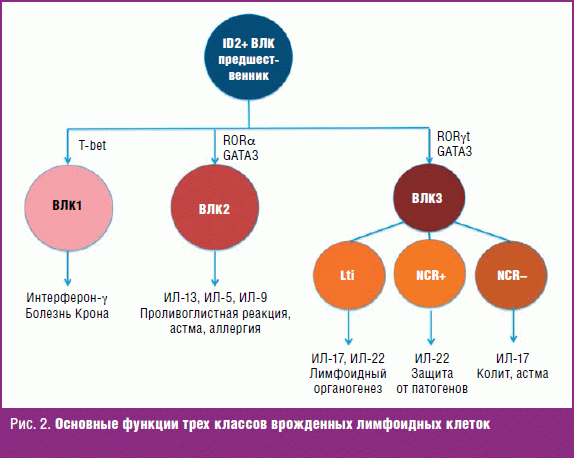
ВЛК имеют общего предшественника, охарактеризованного как клетка, экспрессирующая транскрипторный фактор — ингибитор ДНК-связывающего белка (inhibitor of DNA binding 2, ID2).
ВЛК1 специализируются на ИФН-γ и ФНО-α (защищают слизистую от внутриклеточных паразитов и вирусов); ВЛК2 синтезируют ИЛ-4, ИЛ-5, ИЛ-9 и ИЛ-13 (защита слизистой от многоклеточных паразитов, участие в астме и аллергии); ВЛК3 — преимущественно ФНО-α, интерлейкин-17α и ИЛ-22 (быстро отвечают на инфекцию грибами и внеклеточными бактериями) [54].
ВЛК презентируют Т-лимфоцитам пептидные фрагменты антигенов бактериального происхождения в комплексе с молекулами MHC II класса. Но по причине того, что на мембране ILC-клеток нет ко-стимулирующих молекул, эта презентация носит толерогенный, а не активирующий характер. То есть ВЛК учат Т-лимфоциты «не трогать» симбиотическую флору [55]. Если удастся создать метод генетической модификации ВЛК пациентов так, чтобы усилить у них толерогенную функцию, то может появиться новый метод лечения пациентов в тех популяциях, где ВЛК играют доминирующую роль.
Изменение иммунного ответа под влиянием микробиоты может иметь контактный и дистанционный механизм через Т-регуляторные клетки (Treg), рецепторы врожденного иммунитета (TLR), метаболиты и т. д. [40]. Кишечные бактерии могут модулировать иммунный ответ, непосредственно воздействуя на Tregs через TLR. Комменсальная микробиота способствует дифференцировке индуцированных Tregs (iTreg) из наивных CD4+ T-клеток с помощью независимого механизма первичного ответа TLR и миелоидной дифференцировки TLR и MyD88. Короткоцепочечные жирные кислоты (КЦЖК, или Short-chain fatty acids, SCFA) действуют на Т-клетки через рецептор, связанный с G-белком (GPR43), и защищают макроорганизм от воспаления слизистой кишечника путем увеличения Treg. КЦЖК также способствуют генерации клеток Treg кишечника из наивных CD4+ T-клеток с помощью собственных эпигенетических механизмов Т-клеток. Диета с высоким содержанием клетчатки защищает дыхательные пути от аллергического воспаления, изменяя состав биоты, что приводит к увеличению SCFA. Резидентная микробиота может способствовать развитию Th17-зависимого типа воспаления (показано на мышиной модели астмы, вызванной овальбумином).
Перспективы развития вмешательств при астме и аллергии на основе коррекции микробиоты
Необходим дальнейший анализ влияния микробиомов на фенотипы астмы и аллергии и выявление оптимальных «стратегий, специфичных для фенотипа» (селективное таргетирирование). Полезными могут быть подходы, основанные на манипуляции микробиотой, поскольку средствами традиционной фармакологии можно в лучшем случае подавить иммунитет, но практически невозможно заниматься его регулировкой. Возможное влияние на иммунорегуляцию связывают с пробиотиками, симбиотиками, экобиотиками, синтетическими биотиками или «живыми лекарствами» (генетически модифицированными бактериями для выполнения определенной задачи), трансплантацией фекальной микробиоты, пересадкой редуцированной микрофлоры.
Является ли гипотеза использования пробиотиков при астме и аллергии перспективным направлением? Лейтмотивом исследований с пробиотиками являются доказательства их влияния на иммуномодуляцию. Есть доказательства эффективности пробиотиков in vitro, на животных моделях [56], но индустрия клинических исследований сегодня не успевает за индустрией пробиотиков.
Из-за значительной неоднородности в дизайне исследований, размеров выборки, возраста сбора образцов для изучения микробиоты, методах анализа кишечного микробиома и географического положения в настоящее время нет положительных рекомендаций от научных обществ по использованию пре- или пробиотиков для лечения пищевой аллергии, других аллергических проявлений. Исследования в этой области продолжаются, и некоторые из них показывают возможные преимущества ПБ, которые, по-видимому, практически не имеют риска. Хотя эти исследования предполагают, что использование ПБ может быть многообещающим для профилактики или лечения аллергии, необходимы дополнительные доказательства для определения его эффективности, оптимальной дозировки и штаммов, необходимых для лечения [57].
Cовременные данные не подтверждают пользы рутинного использования ПБ в качестве вмешательства для предотвращения любой формы аллергического заболевания, за исключением группы детей с высоким риском экземы — в соответствии с рекомендацией Всемирной организации по аллергии (World Allergy Organization, WAO) [58].
Оптимальные штаммы, дозы, сроки и продолжительность введения ПБ остаются неизвестными. Тем не менее исследования в этой области продолжаются и позволяют надеяться, что можно прийти к лучшему пониманию роли про-, пре- и синбиотиков в профилактике или лечении аллергии и астмы.
Заключение
В ходе реализации крупных проектов, связанных с микробиомом человека (HMP), произошло радикальное изменение представлений о сложных взаимодействиях организма человека и микрофлоры (микробиоты).
Установлена важная роль микробиоты дыхательных путей (вместе с кишечной микробиотой) в сохранении респираторного здоровья и развитии аллергии/астмы. Секвенирование гена рРНК 16S для идентификации микробиоты выявило наличие Proteobacteria, Firmicutes, Actinobacteria, Fusobacterium и Bacteroides в нижних дыхательных путях здоровых субъектов. У пациентов с астмой доминируют протеобактерии с преобладающими видами Haemophilus. Микробиота дыхательных путей является основным игроком в развитии воспаления слизистой оболочки, дифференцировке лимфоидной ткани, регулировании таких адаптивных ответов, как продукция IgA, функция эффекторных и регуляторных Т-клеток. Микробные компоненты также играют важную роль в регуляции врожденных лимфоидных клеток и функций эпителиальных барьеров. Для формирования иммунной системы важна микробиота кишечника, которая может иметь влияние на респираторную микробиоту и ее функцию в легких через ось «кишечник–легкое».
Изменения в кишечной микробиоте или наличие определенных патогенных бактерий в легких в раннем возрасте связаны с повышенным риском развития аллергии. Но несмотря на стремительный рост исследований в этом направлении остаются неясными механизмы, с помощью которых ось «кишечник–легкое» формирует врожденной и приобретенный иммунный ответ. Предстоит детально изучить, как на микробиоту легкого воздействует окружающая среда, диета, инфекции ВДП у здоровых и у пациентов с астмой, детализировать влияние на микробиоту противоастматических препаратов (длительно действующих бета-агонистов, длительно действующих антихолинергиков). Необходимо уточнить, какую роль играют антибиотики при респираторной патологии и могут ли антибиотики, применяемые при респираторных инфекциях, восстановить нарушенный баланс микробиоты легких при астме. Пока неясны механизмы, через которые микробный дисбиоз приводит к утяжелению течения астмы. Отношения между микробиомом и иммунитетом слизистой оболочки могут быть двунаправленными, то есть конкретные виды комменсальных бактерий могут играть как защитную, так и патогенную роль в развитии аллергии/астмы. Доклинические исследования показали, что изменение микробиоты может модулировать глобальный иммунный ответ хозяина, тем самым уменьшая сенсибилизацию и аллергическое воспаление. Ряд исследований, выполненных на экспериментальных моделях и в клинических условиях, показывает, что эффективная стратегия профилактики и лечения аллергии/астмы может быть связана с воздействием на микробиоту, модуляцию ее состава. Некоторые авторы приводят убедительные обоснования для использования перорального введения пробиотиков/пребиотиков в качестве дополнительных лекарств при аллергии/астме, но имеющиеся данные в этой области недостаточны, чтобы рекомендовать их в рутинной практике.
Иммунная система слизистых находится в постоянной связи с огромным разнообразием микробов, присутствующих в организме человека. Открытие новых молекулярных механизмов, обеспечивающих связь между хозяином и микробами, дает нам ключ к пониманию роли микробиома в сохранении респираторного здоровья.
Литература
- Bunyavanich S., Schadt E. E. Systems biology of asthma and allergic diseases: a multiscale approach // J Allergy Clin Immunol. 2015 Jan; 135 (1): 31–42. DOI: 10.1016/j.jaci.2014.10.015.
- EAACI White Book, on Research, Innovation and Quality Care / Published by the European Academy of Allergy and Clinical Immunology. 2018.
- Kary Mullis. The Polymerase Chain Reaction. Nobel Lecture. Dec. 8. 1993. LEX PRIX NOBEL. The Nobel Prizes. Almqvist and Wiksell Int., Stockholm, Sweden.
- Moraes F., Góes A. A decade of human genome project conclusion: Scientific diffusion about our genome knowledge // Biochem Mol Biol Educ. 2016, May 6; 44 (3): 215–223. DOI: 10.1002/bmb.20952.
- Methе B. A., Nelson K. E., Pop M. et al. A framework for human microbiome research // Nature. 2012, Jun 14; 486 (7402): 215–221. DOI: 10.1038/nature11209.
- The Human Micorbiome Project Consortium. Structure, function and diversity of the healthy human microbiome // Nature. 2012, Jun 13; 486 (7402): 207–214. DOI: 10.1038/nature11234.
- Jason L. P., Galeb A. A., Curtis H. The healthy human microbiome // Genome Med. 2016; 8: 51. Published online 2016. Apr. 27. DOI: 10.1186/ s13073–016–0307-y.
- Turnbaugh P. J., Ley R. E., Hamady M., Fraser-Liggett C. M., Knight R., Gordon J. I. The human microbiome project // Nature. 2007; 449: 804–810.
- Human Microbiome Project: Diversity of Human Microbes Greater Than Previously Predicted. ScienceDaily. Retrieved 8 March 2012.
- Human Microbiome Project — Home | NIH Common Fund. commonfund.nih.gov. Retrieved 2018–04–15.
- Handelsman J. Metagenomics: application of genomics to uncultured microorganisms // Microbiol Mol Biol Rev. 2004; 68: 669–685.
- Handelsman J., Rondon M. R., Brady S. F., Clardy J., Goodman R. M. Molecular biological access to the chemistry of unknown soil microbes: a new frontier for natural products // Chem Biol. 1998, Oct; 5 (10): 245–249.
- National Research Council. Committee on Metagenomics C, Functional A. New science of metagenomics: revealing the secrets of our microbial planet. Washington, DC: National Academies Press; 2007.
- http://www.metahit.eu/.
- http://www.metagenome.ru/.
- Огородова Л. М., Говорун В. М., Федосенко С. В., Карнаушкина М. А., Салтыкова И. В., Алексеев Д. Г., Попенко А. С. Особенности микробиоты кишечника у больных хронической обструктивной болезнью легких // Бюллетень сибирской медицины. 2014; 13 (5): 55–61.
- Федосенко С. В., Огородова Л. М., Карнаушкина М. А., Куликов Е. С., Деев И. А., & Кириллова Н. А. Роль сообщества микроорганизмов дыхательных путей в патогенезе хронической обструктивной болезни легких // Бюллетень сибирской медицины. 2014; 13 (1): 153–160.
- Filion M. Quantitative Real-time PCR in Applied Microbiology. Caister Academic Press: Canada, 2012. 242 р.
- Qin J., Li R., Raes J. et al. A human gut microbial gene catalogue established by metagenomic sequencing // Nature. 2010; 464: 59–65.
- Backhed F., Ding H., Wang T. et al. The gut microbiota as an environmental factor that regulates fat storage // Proc Natl Acad Sci USA. 2004, Nov 2; 101 (44): 15718–15723.
- Резник Н. Л. Внутренний мир // Химия и жизнь. 2013; 8: 8–12.
- Marsh R. L., Nelson M. T., Pope C. E. et al. How low can we go? The implications of low bacterial load in respiratory microbiota studies // Pneumonia (Nathan). 2018, Jul 5; 10: 7. DOI: 10.1186/s41479–018–0051–8.
- Giavina-Bianchi P., Aun M. V., Takejima P. et al. United airway disease: current perspectives // J Asthma Allergy. 2016, May 11; 9: 93–100. DOI: 10.2147/JAA.S81541.
- Bousquet J., Khaltaev N., Cruz A. A. et al. Allergic rhinitis and its impact on asthma (ARIA) 2008 // Allergy. 2008; 63 (86): 8–16.
- Захарова И. Н., Касьянова А. Н., Климов Л. П. и др. Микробиом респираторного тракта: что известно сегодня? // Педиатрия. Прил.к журн. Consilium medicum). 2018; 4: 10–17. DOI:10.26442/24138460.2018.4.180129).
- Salzano F. A., Marino L., Salzano G. et al. Microbiota Composition and the Integration of Exogenous and Endogenous Signals in Reactive Nasal Inflammation // J Immunol Res. 2018, Jun 3; 2018: 2724951. DOI: 10.1155/2018/2724951.
- Stokholm J., Blaser M. J., Thorsen J. et al. Maturation of the gut microbiome and risk of asthma in childhood // Nat Commun. 2018 Jan 10; 9 (1): 141. DOI: 10.1038/s41467–017–02573–2.
- Lambrecht B. N., Hammad H. The immunology of the allergy epidemic and the hygiene hypothesis // Nat Immunol. 2017 Sep 19; 18 (10): 1076–1083. DOI: 10.1038/ni.3829.
- Hilty M., Burke C., Pedro H. et al. Disordered microbial communities in asthmatic airways // PLoS One. 2010; 5: e8578.
- Bassis C. M., Tang A. L., Young V. B., Pynnonen M. A. The nasal cavity microbiota of healthy adults // Microbiome 2014. Aug 11; 2: 27. DOI: 10.1186/2049–2618–2–27.
- Chalermwatanachai T., Velásquez L. C., Bachert C. The microbiome of the upper airways: focus on chronic rhinosinusitis // World Allergy Organ J. 2015, Jan 27; 8 (1): 3. DOI: 10.1186/s40413–014–0048–6.
- Fazlollahi M., Lee T. D., Andrade J. et al. The nasal microbiome in asthma // J Allergy Clin Immunol. 2018 Sep; 142 (3): 834–843. DOI: 10.1016/j.jaci.2018.02.020.
- Charlson E. S. Bittinger K., Haas A. R. et al. Topographical continuity of bacterial populations in the healthy human respiratory tract // Am J Respir Crit Care Med. 2011, Oct 15; 184 (8): 957–63.
- Teo S. M., Mok D., Pham K. et al. The infant nasopharyngeal microbiome impacts severity of lower respiratory infection and risk of asthma development // Cell Host Microbe. 2015, May 13; 17(5): 704–15. DOI: 10.1016/j.chom.2015.03.008.
- Anderson M., Stokken J., Sanford T. et al. A systematic review of the sinonasal microbiome in chronic rhinosinusitis // Am J Rhinol Allergy. 2016, May; 30 (3): 161–166. DOI: 10.2500/ajra.2016.30.4320.
- Venkataraman A., Bassis C. M., Beck J. M. et al. Application of a neutral community model to assess structuring of the human lung microbiome // MBio. 2015; 6: e02284–14.
- Bassis C. M., Erb-Downward J. R., Dickson R. P. et al. Analysis of the upper respiratory tract microbiotas as the source of the lung and gastric microbiotas in healthy individuals // MBio. 2015, Mar 3; 6 (2): e00037. DOI: 10.1128/mBio.00037–15.
- Gleeson K., Eggli D. F., Maxwell S. L. Quantitative aspiration during sleep in normal subjects // Chest. 1997; 111: 1266–72.
- Segal L. N., Alekseyenko A. V., Clemente J. C. et al. Enrichment of lung microbiome with supraglottic taxa is associated with increased pulmonary inflammation // Microbiome. 2013; 1: 19.
- Sokolowska M., Frei R., Lunjani N., Akdis C. A., O’Mahony L. Microbiome and asthma // Asthma Res Pract. 2018, Jan 5; 4: 1. DOI: 10.1186/s40733–017–0037-y.
- Huang Y. J., Nariya S., Harris J. M. et al. The airway microbiome in patients with severe asthma: associations with disease features and severity // J Allergy Clin Immunol. 2015; 136: 874–884. DOI: 10.1016/j.jaci.2015.05.044
- Carr T. F., Kraft M. Chronic infection and severe asthma // Immunol Allergy Clin N Am. 2016; 36: 483–502. DOI: 10.1016/j.iac.2016.03.010;
- Carr T. F., Alkatib R., Kraft M. Microbiome in Mechanisms of Asthma // Clin Chest Med. 2019, Mar; 40 (1): 87–96. DOI: 10.1016/j.ccm.2018.10.006.
- Goleva E., Jackson L. P., Harris J. K. et al. The effects of airway microbiome on corticosteroid responsiveness in asthma // Am J Respir Crit Care Med. 2013; 188: 1193–1201.
- Durack J., Lynch S. V., Nariya S. et al. Features of the bronchial bacterial microbiome associated with atopy, asthma, and responsiveness to inhaled corticosteroid treatment // J Allergy Clin Immunol. 2017; 140: 63–75, DOI: 10.1016/j.jaci.2016.08.055.
- Turturice B. A., McGee H. S., Oliver B. et al. Atopic asthmatic immune phenotypes associated with airway microbiota and airway obstruction // PLoS One. 2017, Oct 20; 12 (10): e0184566.
- Frati F., Salvatori C., Incorvaia C. et al. The Role of the Microbiome in Asthma: The GutLung Axis // Int J Mol Sci. 2018, Dec 30; 20 (1). pii: E123. DOI: 10.3390/ijms20010123.
- Mitchell A. B., Glanville A. R. The Human Respiratory Microbiome: Implications and Impact // Semin Respir Crit Care Med. 2018, Apr; 39 (2): 199–212. DOI: 10.1055/s-0037–1617441.
- Holgate S. T. The airway epithelium is central to the pathogenesis of asthma // Allergol Int. 2008, Mar; 57 (1): 1–10.
- Izuhara K., Nunomura S., Nanri Y. et al. Periostin in inflammation and allergy // Cell Mol Life Sci. 2017 Dec; 74 (23): 4293–4303. DOI: 10.1007/s00018–017–2648–0.
- Doherty T. A. At the bench: understanding group 2 innate lymphoid cells in disease // J Leukoc Biol. 2015, Mar; 97 (3): 455–467.
- Diefenbach A., Colonna M., Koyasu S. Development, Differentiation, and Diversity of Innate Lymphoid Cells // Immunity. 2014, Sep 18; 41(3): 354–365. DOI: 10.1016/j.immuni.2014.09.005.
- Montaldo E., Vacca P., Vitale C. et al. Human innate lymphoid cells // Immunology Letters. 2016; 179: 2–8.
- Yazdani R., Sharifi M., Shirvan A. S. et al. Characteristics of innate lymphoid cells (ILCs) and their role in immunological disorders (an update) // Cell Immunol. 2015, Nov-Dec; 298 (1–2): 66–76.
- Hepworth M. R., Monticelli L. A., Fung T. C. et al. Innate lymphoid cells regulate CD4+ T-cell responses to intestinal commensal bacteria // Nature. 2013, Jun 6; 498 (7452): 113–7. DOI: 10.1038/nature12240.
- Астафьева Н. Г., Гамова И. В., Удовиченко Е. Н., Перфилова И. А., Кобзев Д. Ю., Михайлова И. Э. Иммунная система слизистых: возможности регуляторного действия пробиотиков // РАЖ. 2015; 5: 17–30.
- Wang H. T., Anvari S., Anagnostou K. The Role of Probiotics in Preventing Allergic Disease // Children (Basel). 2019, Feb 5; 6 (2). pii: E24. DOI: 10.3390/children6020024.
- Fiocchi A., Pawankar R., Cuello-Garcia C. A. et al. World Allergy Organization-McMaster University Guidelines for Allergic Disease Prevention (GLAD-P): Probiotics // World Allergy Organ J. 2015, Jan 27; 8 (1): 4.
Д. Ю. Кобзев**, доктор медицинских наук, профессор
И. В. Гамова*, кандидат медицинских наук
И. А. Перфилова*, кандидат медицинских наук
Е. Н. Удовиченко*, кандидат медицинских наук
Л. В. Скучаева*, кандидат медицинских наук
И. Э. Михайлова*
* ФГБОУ ВО СГМУ им. В. И. Разумовского МЗ РФ, Саратов
** Лидс Тринити университет, Лидс, Великобритания
1 Контактная информация: astang@mail.ru
Роль микробиома дыхательных путей в респираторном здоровье (часть 2)/ Н. Г. Астафьева, Д. Ю. Кобзев, И. В. Гамова, И. А. Перфилова, Е. Н. Удовиченко, Л. В. Скучаева, И. Э. Михайлова
Для цитирования: Лечащий врач № 5/2019; Номера страниц в выпуске: 88-92
Теги: микробиом человека, воспаление, бронхиальная обструкция
Купить номер с этой статьей в pdf