Оригинальная статья: Mu Q. et al. Leaky gut as a danger signal for autoimmune diseases //Frontiers in immunology. – 2017. – Т. 8. – С. 598.
Авторы: Qinghui Mu, Jay Kirby, Christopher M. Reilly, Xin M. Luo
Эпителиальная выстилка кишечника выполняет барьерную функцию, предотвращая попадание люминального содержимого (из просвета кишки) в ткани и органы. Нарушение барьерной функции кишечного эпителия приводит к проникновения через стенку кишечника токсинов, пищевых антигенов и микроорганизмов, что лежит в основе патогенеза различных воспалительных и аутоиммунных заболеваний. В литературе появляется все больше данных о ключевой роли микрофлоры кишечника в поддержании нормального функционирования эпителиального барьера и эффективности пробиотиков в предотвращении синдрома повышенной эпителиальной проницаемости, в том числе посредством усиления экспрессии белков плотных контактов. С другой стороны, патогенные бактерии и их метаболиты могут нарушать структуру плотных контактов, способствуя нарушению барьерной функции и развитию аутоиммунных заболеваний. Таким образом, регуляция гомеостаза кишечной микрофлоры является эффективным механизмом защиты кишечного барьера и регуляции эпителиальной проницаемости, и может рассматриваться как потенциальное средство предотвращения и облегчения симптомов аутоиммунных заболеваний.
Ключевые слова: синдром повышенной эпителиальной проницаемости, кишечная микрофлора, бактериальная транслокация, пробиотики, аутоиммунные заболевания
Слизистая оболочка кишечника играет важную роль в пищеварении и поддержании гомеостаза (1). Помимо питательных веществ, абсорбируемых через слизистую оболочку, через эпителиальную выстилку могут проникать различные микроорганизмы и высокомолекулярные вещества, включая пищевые антигены, токсины и бактериальные метаболиты. Для предотвращения проникновения внешних антигенов через стенку желудочно-кишечного тракта (ЖКТ), необходимо постоянное поддержание нормального функционирования эпителиального барьера кишечника. В тонком кишечнике защитный барьер представлен монослоем специализированных эпителиальных клеток, связанных друг с другом белками плотных контактов, а также различными гуморальными факторами, включая муцины, иммуноглобулины и цитокины. Нарушение нормального функционирования звеньев этой системы может приводить к повышению проницаемости эпителиальной выстилки, и, как следствие, проникновению внешних агентов через межклеточные барьеры, что оказывает патологическое воздействие на работу кишечника и организма в целом. Нарушение барьерной функции кишечника лежит в основе патогенеза различных аутоиммунных и воспалительных заболеваний, включая синдром раздраженного кишечника, целиакию, аутоиммунный гепатит, сахарный диабет 1 типа, рассеянный склероз и системную красную волчанку (2–6).
На функционирование кишечного барьера оказывают влияние различные факторы, включая качественный и количественный состав кишечной микрофлоры, пищевые антигены и алкоголь. Нарушение эпителиальной проницаемости характерно не только для людей, но и для других млекопитающих, что позволяет исследовать механизмы нарушения барьерной функции кишечника на животных моделях (7, 8).
В последние годы все больший интерес вызывает изучение роли микрофлоры кишечника в патогенезе синдрома повышенной эпителиальной проницаемости и регуляции барьерной функции кишечника (9). В настоящем обзоре представлены последние данные о механизмах и патогенетических факторах, способствующих повышению проницаемости кишечной стенки и нарушению барьерной функции кишечного эпителия, и о связи между кишечной проницаемостью и наиболее распространенными аутоиммунными заболеваниями.
Кишечный барьер
В кишечник человека попадает большое количество внешних агентов, включая разнообразные микроорганизмы, токсины и антигены. Нарушение барьерной функции кишечной стенки облегчает проникновение этих агентов через эпителиальную выстилку кишечника в кровоток и лимфатическую систему. Физические, биохимические и иммунологические особенности барьерной системы тонкого кишечника позволяют предотвратить проникновение большинства патогенов из полости кишечника в ткани и органы (Рисунок 1).
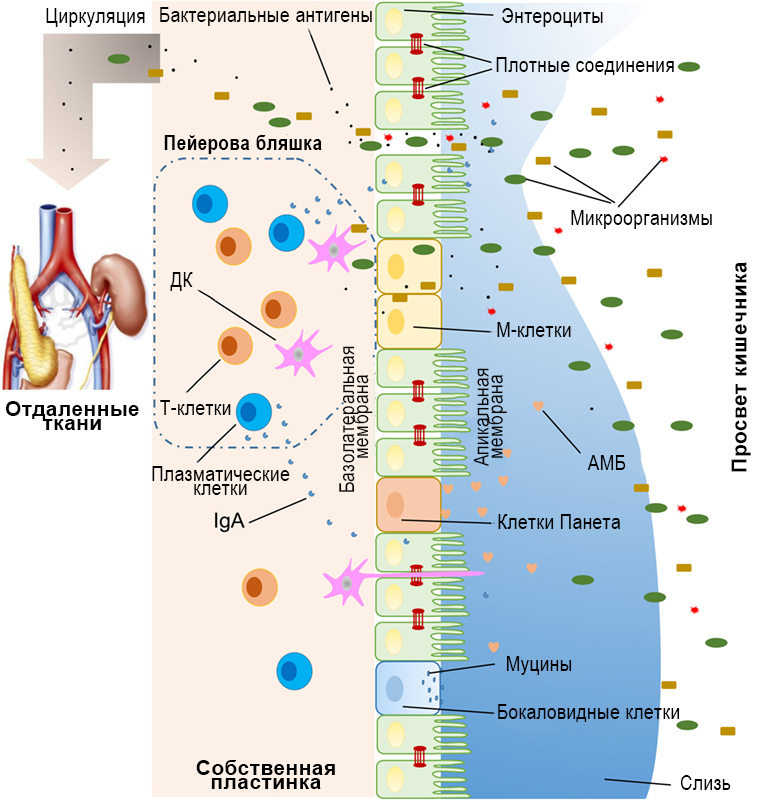
Рисунок 1. Схематическое изображение кишечного барьера, в т.ч. физического (эпителий, плотные контакты, слизь, комменсальные бактерии), биохимического (АМП) и иммунологического (лимфоциты и IgA).
Физический барьер
Основу физического барьера кишечника составляет эпителиальный слой, общая площадь которого превышает 400 кв. м (1) (Рисунок 1). Кишечный эпителий состоит из семи функциональных подтипов кишечных эпителиальных клеток: энтероцитов, бокаловидных клеток, клеток Панета или ацидофильных энтероцитов, эндокриноцитов, микроскладчатых клеток (М-клеток), чашеобразных и пучковых клеток, хотя функция последних двух типов до сих пор малоизучена (10). Энтероциты – абсорбирующие клетки, составляющие основную массу клеток эпителия кишечника и до 90 % клеток ворсинок и кишечных крипт. Функция энтероцитов не ограничивается всасыванием питательных веществ. Появляется все больше сведений об их роли в регуляции количества грамположительных бактерий посредством экспрессии антимикробного белка (АМП) RegIIIγ семейства регенераторных Reg-антимикробных пептидов (11–13). Все эпителиальные клетки кишечника дифференцируются из эпителиальных стволовых клеток линии Lgr5+, расположенных в основании кишечных крипт (14). У млекопитающих клетки кишечного эпителия обновляются каждые 3–5 дней (10, 15), за исключением клеток Пенета, цикл жизни которых составляет до 2-х месяцев.
Эпителиальная выстилка кишечной стенки непрерывна. Механическое соединение клеток обеспечивается плотными контактами (16), через которые осуществляется парацеллюлярный транспорт веществ из полости кишечника в кровоток и лимфу. Известно более 40 белков, входящих в эти комплексы, среди которых наиболее хорошо изучены окклюдины, клаудины, молекулы клеточной адгезии семейства JAMs (JAM-A) и трицеллюлин (17). Увеличение парацеллюлярного транспорта вследствие патологических процессов в кишечнике может приводить к проникновению высокомолекулярных соединений из интерстиция в кровоток.
Слизистый слой эпителиальной выстилки кишечника представлен двумя слоями – внутренним и внешним, ограничивающими эпителий от кишечных бактерий и продуктов их метаболизма. Основным компонентом слизи в тонком и толстом кишечнике является гликозилированный гелеобразующий муцин второго типа (MUC2), секретируемый бокаловидными клетками. Помимо этого, слизистый слой содержит различные активные молекулы, в частности IgA, ферменты и белки, включая лактоферрин (18). Бокаловидные клетки продуцируют не только муцин MUC2, но и другие важные компоненты слизи – ZG16, AGR2, FCGBP, CLCA1 и TFF3 (19, 20). У мышей с нокаутом гена Muc2 развивается спонтанный колит, что подчеркивает защитную функцию MUC2 (21). Помимо гельобразующих муцинов, в слизистом слое кишечника присутствуют трансмембранные муцины, экспрессируемые преимущественно энтероцитами и находящиеся вблизи апикальной поверхности эпителиальных клеток (20).
Комменсальные бактерии способствуют поддержанию барьерной функции кишечного эпителия (22). Во-первых, они предотвращают колонизацию слизистой оболочки патогенными микроорганизмами, создавая конкурентную среду и занимая поверхность эпителиальной выстилки, и высвобождая антимикробные соединения (23, 24). Во-вторых, бактерии кишечной микрофлоры участвуют в регуляции переваривания и абсорбции питательных веществ, служащих источником энергии для эпителиальных клеток (25). Короткоцепочечные жирные кислоты, синтезируемые кишечными бактериями, служат основным источником энергии для клеток слизистой толстого кишечника (26). Таким образом, эпителиальные клетки кишечника, слизистый слой и кишечная микробиота составляют физический барьер, препятствующий проникновению экзогенных факторов из содержимого кишечника во внутреннюю среду организма.
Биохимический барьер
Усилению барьерных функций слизистой оболочки кишечника способствуют молекулы, обладающие антимикробным действием, в частности желчные кислоты и антимикробные пептиды (АМП) (27, 28) (Рисунок 1). Эти соединения усиливают барьерную функцию эпителия и препятствуют прямому контакту люминального содержимого с тканями и органами.
Количество микроорганизмов значительно увеличивается по мере продвижения от проксимальных отделов ЖКТ, где обнаруживается лишь небольшое количество бактерий (29), к дистальным (30). АМП, продуцируемые представителями кишечной микрофлоры, включают α- и β-дефенсины, лектин С, кателицидины, лизоцим и кишечную щелочную фосфатазу (КЩФ) (27). Антимикробные свойства этих пептидов подробно обсуждаются в работе Kopp с соавторами (31). В эпителиальной выстилке кишечника антибактериальные пептиды секретируются преимущественно клетками Панета.
Иммунологический барьер
Иммунологический барьер кишечника представлен одиночными лимфоидными фолликулами и пейеровыми бляшками – периферическими скоплениями лимфоидных клеток, расположенными в собственной пластинке слизистой оболочки тонкой кишки. Внутри фолликулов находятся различные иммунные элементы, включая В- и Т-лимфоциты, дендритные клетки (ДК) и нейтрофилы, секретирующие цитокины и антитела в ответ на проникновение антигена (Рисунок 1). М-клетки участвуют в трансцитозе антигенов из просвета кишечника к подлежащим иммунным клеткам (14).
Бокаловидные клетки участвуют в презентации люминальных антигенов комплексу CD103+ ДК собственной пластинки слизистой оболочки кишечника, образуя антигенные комплексы (goblet cell-associated antigen passages, GAPs) (32, 33). В толстом кишечнике у стерильных мышей или животных, получающих антибиотики, также может происходить спонтанная презентация антигена (34). Это указывает на роль кишечной микрофлоры в захвате антигенов и образовании GAPs (35). Кроме того, бокаловидные клетки и GAPs способны подавлять транслокацию патогенных бактерий из просвета кишечника во внутреннюю среду организма (36). Антигены также могут связываться и распознаваться субэпителиальными мононуклеарными фагоцитами (37, 38), экспрессирующими на поверхности рецептор фракталкина CX3CR1. Распознавание антигенов происходит за счет трансэпителиальных дендритов (39, 40), образование которых регулируется CX3CR1+ макрофагами и экспрессией CX3CL1 некоторыми клетками кишечного эпителия (41, 42).
Секреторные IgA (SIgA), еще один компонент иммунного барьера кишечника, продуцируются плазмоцитами (в организме взрослого человека – 50 мг/кг SIgA
ежедневно) и локализуются преимущественно в собственной пластинке слизистой оболочки кишечника. Считается, что SIgA способны взаимодействовать с комменсальными бактериями кишечника, опосредуя формирование бактериальной биопленки. Важной особенностью SIgA является их устойчивость к действию кишечных протеаз, что обеспечивает поддержание защитой функции даже в условиях агрессивной среды кишечника (43). SIgA могут проникать через эпителиальную выстилку в просвет кишечника, связывать антигены и доставлять их к иммунным клеткам лимфоидной ткани.
Важную роль в поддержании нормального функционирования барьерной функции кишечника также играют различные химические факторы и нервная регуляция.
Микрофлора кишечника и кишечный барьер
Распознавание бактерий в кишечнике опосредовано рецепторами PRR (образраспознающие рецепторы, Pattern-Recognition Receptors), распознающими патоген-ассоциированные молекулярные паттерны (РАМР), включая toll-подобные рецепторы (TLR) и нуклеотид-связывающие домен-рецепторы олигомеризации (NOD-подобные рецепторы, NLR). Взаимодействие между бактериями кишечника и клетками хозяина во многом зависит от распознавания микробно-ассоциированных молекулярных паттернов образраспознающими рецепторами PRR, экспрессируемыми на поверхности иммунных и неиммунных клеток.
Продукты жизнедеятельности бактерий также способны нарушать барьерную функцию кишечной стенки и изменять проницаемость кишечного барьера. Одним из последствий нарушения целостности кишечного барьера и повышения эпителиальной проницаемости является бактериальная транслокация, которая сопровождается проникновением бактерий и их метаболитов через эпителиальный слой, вызывая системное воспаление и повреждение тканей (Рисунок 1) (44).
Результаты экспериментальных исследований на стерильных животных указывают на критическую роль кишечной микрофлоры в развитии и поддержании барьерной функции кишечника. Отсутствие бактериальной флоры у стерилизованных мышей приводит к снижению толщины слизистого слоя кишечника (45–48). Качественный и количественный состав кишечной микрофлоры вносит значительный вклад в поддержание целостности слизисто-эпителиального барьера кишечника за счет регуляции продукции слизи бокаловидными клетками (49, 50). Снижение толщины слизистого слоя облегчает проникновение микроорганизмов в субэпителиальное пространство, вызывая воспалительную реакцию и ассоциированные заболевания, в частности колит (46, 51).
Комменсальные бактерии и бактериальные продукты, например, липополисахариды (ЛПС) и пептидогликан, способствуют восстановлению слизистого слоя кишечника (46, 47). Все вместе, комменсальные бактерии и слизистый слой играют основополагающую роль в поддержании кишечного гомеостаза (48). Разнообразные АМП, секретируемые в слизистом слое и активные в отношении патогенных микроорганизмов, позволяют угнетать их активность и регулировать колонизацию кишечника комменсальными бактериями. Секреция некоторых АМП реципрокно регулируется кишечной микробиотой и/или ее бактериальными продуктами. Например, активность антимикробного пептида RegIIIγ, способствующего отделению комменсальных бактерий от кишечного эпителия (11), снижена у пациентов на фоне частого употребления алкоголя и у мышей, получающих этанол (52, 53). Прием пребиотиков, в частности Lactobacilli и Bifidobacteria, позволяет восстановить активность RegIIIγ и стимулировать рост нормальной микрофлоры кишечника (53). Синтез и секреция фактора ангиогенеза Ang4, обладающего антибактериальной активностью, клетками Панета мышей также преимущественно регулируется представителями облигатной кишечной микрофлоры Bacteroides thetaiotaomicron (54).
Кишечная микробиота может взаимодействовать с рядом АМП, в частности кишечной щелочной фосфатазой КЩФ, продуцируемой эпителиальными клетками кишечника и связанной с клеточной мембраной. При секреции в просвет кишечника активность КЩФ сохраняется (55, 56). При недостатке щелочной фосфатазы у нокаутных мышей наблюдается нарушение естественного гомеостаза кишечной микробиоты, в частности снижение популяции Lactobacillaceae, по сравнению с особями дикого типа (57, 58). Повышение активности КЩФ избирательно повышает количество пробиотических бактерий (напр., Bifidobacterium), подавляющих патогенный рост ЛПС-продуцирующих микроорганизмов (напр., Escherichia coli) (59). Способность КЩФ инактивировать ЛПС in vivo обуславливает ключевую роль этого фермента в предотвращении проникновения бактериальных ЛПС в кишечную стенку (60, 61).
Кишечная микробиота также способна регулировать экспрессию КЩФ. У стерильных рыб Danio rerio, добавление бактериальных ЛПС или колонизация кишечника комменсальными микроорганизмами в значительной степени повышает экспрессию КЩФ (62). Влияние КЩФ на усиление барьерной функции кишечного эпителия также обусловлено способностью этого фермента регулировать продукцию белков плотных контактов и увеличивать экспрессию ZO-1, ZO-2 и окклюдина (63). Рядом исследований продемонстрирована роль различных АМП в реципрокной регуляции состава кишечной микрофлоры (64, 65).
Источником образования новых эпителиальных клеток кишечника, образующих непрерывный монослой, служат стволовые клетки, расположенные на базальной части кишечных крипт и экспрессирующие TLR4 рецепторы к ЛПС. Непрерывное и быстрое обновление клеток кишечного эпителия играет ключевую роль в поддержании барьерной функции эпителиальной выстилки (66). Около 10% транскрипции генов, связанных с иммунной функцией, клеточной пролиферацией и метаболизмом в эпителиальных клетках кишечника регулируется бактериями кишечника (67). Снижение скорости пролиферации эпителиальных клеток у стерильных мышей, лишенных бактериальной флоры, или животных, получающих антибиотики, указывает на важную роль кишечной микрофлоры в обновлении клеток эпителия (68, 69). ЛПС E. coli способны дозозависимо индуцировать отслоение клеток (70, 71). Модуляция ЛПС-индуцируемого отслоения эпителиальных клеток экзополисахаридами Bifidobacterium breve происходит через MyD88-зависимый сигнальный путь (70).
Активация TLR4, расположенных на мембране стволовых клеток кишечных крипт, бактериальными ЛПС подавляет пролиферацию и индуцирует апоптоз Lgr5+ стволовых клеток кишечника. В ответ на селективную инактивацию TLR4 стволовых клеток кишечника снижается способность ЛПС подавлять обновление клеток эпителия (72). Этот процесс опосредован действием p53-зависимого модулятора апоптоза PUMA, на что указывается сохранение активности TLR4 в отсутствие этого белка. Архитектура кишечных крипт защищает стволовые клетки от негативного влияния других бактериальных метаболитов, например, бутирата, способного, наряду с ЛПС, подавлять клеточную пролиферацию (73). Пробиотические микроорганизмы, в частности Lactobacillus rhamnosus (75–78), Streptococcus thermophilus (79), Lactobacillus reuteri (80) и Bifidobacterium infantis (81) могут влиять на экспрессию белков плотных контактов, служащих основным барьерным механизмом, препятствующим проникновению патогенов через кишечный эпителий.
Механизмы синдрома повышенной эпителиальной проницаемости
Нарушение барьерной функции эпителиальной выстилки кишечника и изменение качественного и количественного состава кишечной микрофлоры в ответ на воздействие неблагоприятных факторов (алкоголь, инфекционные заболевания, диетические факторы) может приводить к транслокации антигенов, патогенных микроорганизмов и их токсинов через эпителий, вызывая системное и местное воспаление.
Влияние диетических факторов на эпителиальную проницаемость
Состав кишечной микробиоты и барьерная функция кишечника зависят от диетических предпочтений человека (82, 83).
Положительное влияние витамина D на функцию слизисто-эпителиального барьера кишечника опосредовано его регуляторным воздействием на экспрессию белков плотных контактов ZO-1 и клаудина-1. Инактивация транскрипции рецепторов витамина D (VDR) приводит к развитию колита (84). С другой стороны, повышение экспрессии VDR повышает активность промотора порообразующего клаудина-2, обеспечивающего параклеточный транспорт, сопряженный с повышением проницаемости плотных контактов (85). Таким образом, роль витамина D и VDR в регуляции кишечной проницаемости до сих пор остается предметом дискуссий.
Диета с низким содержанием пищевых волокон провоцирует рост бактерий, нарушающих целостность слизистой оболочки кишечника, в частности Akkermansia muciniphila и Bacteroides caccae (45), что приводит к снижению толщины слизистого слоя. При этом, транскрипция гена Muc2 повышается, что указывает на активацию компенсаторных механизмов. Снижение барьерных функций и толщины слизистого слоя повышают чувствительность к патогенным бактериям, способных провоцировать колит (45).
Диета, богатая насыщенными жирами и сахарами, вызывает снижение популяции Lactobacillus и рост Oscillibacter, что приводит к повышению проницаемости межклеточного барьера в проксимальных отделах толстого кишечника (86) и снижению экспрессии белка плотных контактов ZO-1.
Влияние стрессорных факторов на эпителиальную проницаемость
Проницаемость слизисто-эпителиального барьера кишечника для патогенных микроорганизмов может нарушаться под влиянием стрессорных факторов, в частности при ожоговых поражениях и длительном употреблении алкоголя.
Повышение эпителиальной проницаемости, ассоциированной с ожоговым шоком, опосредовано увеличением активности киназы легких цепей миозина (MLC) (87, 88), приводящим к изменениям состояния актин-миозиновых комплексов и расширению плотных соединений между клетками кишечного эпителия (89–91). Открытие плотных контактов опосредовано перераспределением и перемещением ZO-1, окклюдина и клаудина-1 внутрь клетки, что при ожогах может быть предотвращено добавлением ингибитора фосфорилирования легких цепей миозина (87). Изменения микробиоценоза в результате ожогового поражения приводят к нарушению естественного гомеостаза кишечной микрофлоры, и, как следствие, увеличению количества и перемещению грамотрицательных аэробных условно-патогенных микроорганизмов, в том числе представителей семейства Enterobacteriaceae (88) через стенку толстого кишечника. Вместе с этим наблюдается снижение популяции аэробных и анаэробных бутират-продуцирующих бактерий (92), которое сопровождается снижением концентрации бутирата в фекалиях мышей с ожоговым поражением. Восстановление качественного и количественного состава микробиоты и функции слизистого барьера в ответ на фекальную трансплантацию подчеркивает непосредственное участие патогенных микроорганизмов в патогенезе повышенной проницаемости кишечника при ожоговых поражениях.
Хроническое употребление алкоголя также приводит к нарушению барьерной функции кишечной стенки, вызывая количественное и качественное нарушение естественного гомеостаза кишечной микробиоты, транслокацию ЛПС и алкогольную болезнь печени (АБП). Повреждение эпителиального барьера кишечника, вызванное систематическим употреблением алкоголя, опосредовано повышением секреции фактора некроза опухоли (ФНО) кишечными моноцитами и макрофагами кишечника, а также увеличением уровня энтероцитов, несущих ФНО-рецепторы типа I, что приводит к угнетению активности киназы легких цепей миозина (93). Воздействие антибиотиков на фоне хронического введения алкоголя мышам приводит к снижению экспрессии ФНО и восстановлению проницаемости кишечной стенки до нормальных значений (93), что подчеркивает роль кишечной микробиоты в патогенезе повышенной проницаемости кишечного эпителия, опосредованной действием ФНО. Систематическое употребление алкоголя сопровождается избыточным ростом популяции патогенных микроорганизмов и снижением количества Lactobacillus (53), хотя точный механизм, опосредующий действие алкоголя на качественный и количественный состав кишечной микробиоты до сих пор неизвестен (94). Применение антибиотиков, наоборот, способно снижать бактериальную нагрузку и уменьшать выраженность симптомов АБП (53, 93, 95–97). Увеличение популяции Lactobacillus способствует снижению роста и транслокации патогенных микроорганизмов и восстановлению целостности слизистой оболочки кишечника (53, 94, 98, 99).
Микробная транслокация, особенно транслокация ЛПС, является одним из ключевых патогенетических механизмов АБП, что подтверждается данными об отсутствии симптомов АБП при инактивации TLR4, в норме экспрессируемых всеми типами печеночных клеток (100, 101). Важно отметить, что эндогенным источником этанола также могут служить некоторые кишечные бактерии, в частности E. coli и Weissella confusa, что может лежать в основе патогенеза нарушения барьерной функции кишечного эпителия (102, 103).
Установлена роль инфекционных агентов в изменении свойств кишечного барьера. Так, например, грамотрицательные бактерии Helicobacter pylori способны напрямую взаимодействовать с белками плотных контактов ZO-1, вызывая повышение проницаемости эпителия (104–106). Помимо бактериальных агентов, продемонстрирована роль бактериофагов, в частности фагов против Salmonella enterica, в нарушении кишечного барьера и патогенезе синдрома повышенной проницаемости кишечника, что, вероятно, также опосредовано негативным влиянием на популяцию нормальной микрофлоры кишечника (107).
Таким образом, нарушение барьерной функции кишечного эпителия и транслокация патогенных микроорганизмов и их метаболитов через кишечную стенку, вызванные изменением состава нормальной микрофлоры, вносят значительный вклад в повышение проницаемости эпителиального барьера, активации иммунного ответа, формированию воспалительных изменении? и повреждению тканей.
Синдром повышенной эпителиальной проницаемости и аутоиммунные заболевания
Дисбиоз, повышение бактериальной транслокации и нарушения барьерной функции кишечника могут быть связаны с некоторыми неинфекционными заболеваниями, включая болезнь Альцгеймера, АБП, онкологические и аутоиммунные заболевания.
Аутоиммунные патологии сопровождаются формированием иммунного ответа на собственные антигены организма, что приводит к повреждению клеток, экспрессирующих аутоантигены. Появляется все больше сведений о роли бактериальной транслокации и дисфункции эпителиального барьера кишечника в патогенезе аутоиммунных заболеваний (2–6).
Сахарный диабет 1 типа
Сахарный диабет 1 типа (СД1) – хроническое заболевание эндокринной системы, связанное с аутоиммунным поражением β-клеток островков Лангерганса поджелудочной железы, обуславливающим снижение продукции и возникновение дефицита инсулина (108). Некоторые исследователи считают, что нарушение проницаемости слизисто-эпителиального барьера является скорее следствием, нежели причиной СД1 (109). Однако гораздо больше данных свидетельствует в пользу обратного.
В исследованиях с участием пациентов с СД1 и на животных моделях продемонстрировано, что нарушения барьерной функции кишечника предшествуют началу заболевания (110–112). Кроме того, установлено, что повышение проницаемости кишечного барьера при СД1 ассоциировано с увеличением уровня зонулина – белка, способного связываться с рецепторами плотных контактов и вызывать увеличение межэпителиальных промежутков. Установлено, что изменение уровня зонулина также происходит под действием факторов кишечной микробиоты (113). Применение ингибиторов зонулина приводит к восстановлению барьерной функции кишечника и облегчению течения заболевания у склонных к диабету мышей (114).
Бактериальная транслокация также может способствовать развитию СД1 (115). В модели стрептозотоцин-индуцированного СД1 введение стрептозотоцина способствовало восстановлению гомеостаза кишечной микробиоты по сравнению с плацебо. Кроме того, продемонстрирована способность кишечных бактерий проникать в региональные лимфатические узлы поджелудочной железы, вызывая активацию NOD2 и развитие симптомов СД1 (115). Введение антибиотиков, наоборот, приводило к элиминации бактерий из ткани поджелудочной железы и снижению проявлений заболевания.
Системная красная волчанка
Системная красная волчанка (СКВ) – хроническое аутоиммунное заболевание, при котором наблюдается продукция аутоиммунных антител к тканям собственного организма, что приводит к хроническому воспалению и повреждению тканей и органов (116). У женщин детородного возраста СКВ встречается в 9 раз чаще, чем у мужчин.
Проникновение ЛПС грамотрицательных бактерий через кишечную стенку и их транслокация в ткани играет важную роль в развитии и прогрессировании СКВ (117). Повышение уровня растворимого бактериального маркера транслокации CD14 у пациентов с СКВ указывает на увеличение циркуляции ЛПС и активацию моноцитов (118).
Бактериальные ЛПС – основной лиганд TLR4. Установлено, что активация этих рецепторов инициирует развитие СКВ (119–121). В ответ на стимуляцию TLR4 у мышей наблюдается спонтанное развитие СКВ, в то время как элиминация комменсальной бактериальной флоры антибиотиками позволяет добиться облегчения симптомов заболевания (121), что свидетельствует о важной роли гиперчувствительности TLR4 к бактериальным ЛПС в патогенезе СКВ.
Введение ЛПС позволяет облегчить течение СКВ у мышей дикого типа (C57BL/6 или BALB/c), иммунизированных фосфолипид-связывающим белком (122–124). Инактивация TLR4, напротив, приводит к снижению уровня аутоантител и концентрации IgG в почечных клубочках мышей, предрасположенных к развитию СКВ (125, 126). Таким образом, активация TLR4 бактериальными ЛПС является фактором, инициирующим развитие СКВ.
Помимо ЛПС вклад в развитие СКВ вносит липотейхоевая кислота (ЛТА), компонент стенки грамположительных бактериальных клеток. Показано, что у пациентов с СКВ увеличена экспрессия TLR2, рецептора ЛТА (127). Кроме того, у мышей, предрасположенных с СКВ, активация TLR2 приводит к развитию волчаночного нефрита, в то время как инактивация этого рецептора облегчает симптомы заболевания (125, 128–130). В последних исследованиях также был обнаружен еще один антиген, способный инициировать продукцию аутоантител в результате антигенной мимикрии (131).
Некоторые белки сигнального каскада TLR, в частности MyD88, IRAKs, and IFNα, участвующие в патогенезе СКВ, могут выступать в качестве потенциальных терапевтических мишеней (132). Облегчение симптомов СКВ у мышей линии MRL/lpr с нокаутом гена MyD88 указывает на важную роль взаимодействия между TLR и патогенной микрофлорой в развитии и прогрессировании СКВ (133, 134).
Рядом исследований продемонстрирована роль nod-подобных рецепторов (NLR), в частности компонентов инфламмасом NLRP3 и AIM2, участвующих в распознавании комменсальных бактерий, в патогенезе СКВ (135–137). У мышей линии NZB обнаружено снижение активности NLRP3- и AIM2-инфламмасом, а у мышей линии B6-Faslpr инактивация адаптерного апоптоз-ассоциированного speck-подобного белка, содержащего CARD (ASC), необходимого для образования инфламмасом, приводила к манифестации СКВ (138).
Таким образом, роль TLR и NLR в патогенезе СКВ опосредована способностью этих рецепторов связываться с антигенами представителей патогенной и комменсальной микрофлоры, соответственно, которые могут проникать через кишечную стенку при повышении проницаемости эпителиального барьера.
Регуляция эпителиальной проницаемости как метод терапии
Вклад нарушений барьерной функции и бактериальной транслокации в индукцию воспаления и патогенез различных заболеваний позволяет рассматривать механизмы проницаемости кишечной стенки в качестве потенциальной терапевтической мишени. Например, пребиотики и пробиотики, в частности комменсальные бактерии Bacteroides fragilis, могут быть успешно применены для снижения проницаемости кишечника и защиты эпителиального барьера (139, 140).
Как показывают эксперименты на животных моделях с аутизмом , расстройства аутистического спектра (РАС) сопровождаются дисфункцией эпителиального барьера кишечника, дисбиозом и повышенной проницаемостью кишечной стенки для 4-этилфенилсульфата (4ЭФС), продуцируемого комменсальными бактериями. Введение 4ЭФС здоровым мышам вызывает изменения поведения, характерные для РАС. Введение B. fragilis, наоборот, приводит к снижению транслокации 4EPS и нормализации поведения животных.
Считается, что терапевтическая роль B. fragilis опосредована способностью бактерии нормализовать состав кишечной микрофлоры и повышать барьерную функцию кишечника (140). Кроме того, полисахарид А, метаболит B. fragilis, индуцирует развитие Foxp3+Т-регуляторных клеток (141, 142). B. fragilis и полисахарид А также продемонстрировали высокую эффективность при некоторых воспалительных заболеваниях, включая колит и экспериментальный аутоиммунный энцефаломиелит (141, 143). Тем не менее, для широкого применения B. fragilis в предотвращении и лечении синдрома повышенной эпителиальной проницаемости требуется больше исследований. Поскольку пробиотики в большинстве случаев обладают хорошим профилем безопасности, можно ожидать, что в будущем они будут более широко применяться в терапии аутоиммунных заболеваний.
Литература
1. Peterson LW, Artis D. Intestinal epithelial cells: regulators of barrier function and immune homeostasis. Nat Rev Immunol (2014) 14(3):141–53. doi: 10.1038/nri3608
2. Lin R, Zhou L, Zhang J, Wang B. Abnormal intestinal permeability and microbiota in patients with autoimmune hepatitis. Int J Clin Exp Pathol (2015) 8(5):5153–60.
3. Khaleghi S, Ju JM, Lamba A, Murray JA. The potential utility of tight junction regulation in celiac disease: focus on larazotide acetate. Therap Adv Gastroenterol (2016) 9(1):37–49. doi:10.1177/1756283X15616576
4. Fasano A, Shea-Donohue T. Mechanisms of disease: the role of intestinal barrier function in the pathogenesis of gastrointestinal autoimmune diseases. Nat Clin Pract Gastroenterol Hepatol (2005) 2(9):416–22. doi:10.1038/ncpgasthep0259
5. Tlaskalova-Hogenova H, Stepankova R, Kozakova H, Hudcovic T, Vannucci L, Tuckova L, et al. The role of gut microbiota (commensal bacteria) and the mucosal barrier in the pathogenesis of inflammatory and autoimmune diseases and cancer: contribution of germ-free and gnotobiotic animal models of human diseases. Cell Mol Immunol (2011) 8(2):110–20. doi:10.1038/cmi.2010.67
6. Fasano A. Zonulin, regulation of tight junctions, and autoimmune diseases. Ann N Y Acad Sci (2012) 1258:25–33. doi:10.1111/j.1749-6632.2012.06538.x
7. Jiminez JA, Uwiera TC, Douglas Inglis G, Uwiera RR. Animal models to study acute and chronic intestinal inflammation in mammals. Gut Pathog (2015) 7:29. doi:10.1186/s13099-015-0076-y
8. Brugman S. The zebrafish as a model to study intestinal inflammation. Dev Comp Immunol (2016) 64:82–92. doi:10.1016/j.dci.2016.02.020
9. Hooper LV, Littman DR, Macpherson AJ. Interactions between the microbiota and the immune system. Science (2012) 336(6086):1268–73. doi:10.1126/science.1223490
10. Gerbe F, Legraverend C, Jay P. The intestinal epithelium tuft cells: specification and function. Cell Mol Life Sci (2012) 69(17):2907–17. doi:10.1007/s00018-012-0984-7
11. Vaishnava S, Yamamoto M, Severson KM, Ruhn KA, Yu X, Koren O, et al. The antibacterial lectin RegIIIgamma promotes the spatial segregation of microbiota and host in the intestine. Science (2011) 334(6053):255–8. doi:10.1126/science.1209791
12. Cash HL, Whitham CV, Behrendt CL, Hooper LV. Symbiotic bacteria direct expression of an intestinal bactericidal lectin. Science (2006) 313(5790):1126–30. doi:10.1126/science.1127119
13. Vaishnava S, Behrendt CL, Ismail AS, Eckmann L, Hooper LV. Paneth cells directly sense gut commensals and maintain homeostasis at the intestinal host-microbial interface. Proc Natl Acad Sci U S A (2008) 105(52):20858–63. doi:10.1073/pnas.0808723105
14. Mabbott NA, Donaldson DS, Ohno H, Williams IR, Mahajan A. Microfold (M) cells: important immunosurveillance posts in the intestinal epithelium. Mucosal Immunol (2013) 6(4):666–77. doi:10.1038/mi.2013.30
15. van der Flier LG, Clevers H. Stem cells, self-renewal, and differentiation in the intestinal epithelium. Annu Rev Physiol (2009) 71:241–60. doi:10.1146/annurev.physiol.010908.163145
16. Anderson JM, Van Itallie CM. Physiology and function of the tight junction. Cold Spring Harb Perspect Biol (2009) 1(2):a002584. doi:10.1101/cshperspect.a002584
17. Yamazaki Y, Okawa K, Yano T, Tsukita S, Tsukita S. Optimized proteomic analysis on gels of cell-cell adhering junctional membrane proteins. Biochemistry (2008) 47(19):5378–86. doi:10.1021/bi8002567
18. Singh PK, Parsek MR, Greenberg EP, Welsh MJ. A component of innate immunity prevents bacterial biofilm development. Nature (2002) 417(6888):552–5. doi:10.1038/417552a
19. Hansson GC. Role of mucus layers in gut infection and inflammation. Curr Opin Microbiol (2012) 15(1):57–62. doi:10.1016/j.mib.2011.11.002
20. Pelaseyed T, Bergstrom JH, Gustafsson JK, Ermund A, Birchenough GM, Schutte A, et al. The mucus and mucins of the goblet cells and enterocytes provide the first defense line of the gastrointestinal tract and interact with the immune system. Immunol Rev (2014) 260(1):8–20. doi:10.1111/imr.12182
21. Van der Sluis M, De Koning BA, De Bruijn AC, Velcich A, Meijerink JP, Van Goudoever JB, et al. Muc2-deficient mice spontaneously develop colitis, indicating that MUC2 is critical for colonic protection. Gastroenterology (2006) 131(1):117–29. doi:10.1053/j.gastro.2006.04.020
22. Sekirov I, Russell SL, Antunes LC, Finlay BB. Gut microbiota in health and disease. Physiol Rev (2010) 90(3):859–904. doi:10.1152/physrev.00045.2009
23. Baumler AJ, Sperandio V. Interactions between the microbiota and pathogenic bacteria in the gut. Nature (2016) 535(7610):85–93. doi:10.1038/nature18849
24. Ming L, Zhang Q, Yang L, Huang JA. Comparison of antibacterial effects between antimicrobial peptide and bacteriocins isolated from Lactobacillus plantarum on three common pathogenic bacteria. Int J Clin Exp Med (2015) 8(4):5806–11.
25. Ramakrishna BS. Role of the gut microbiota in human nutrition and metabolism. J Gastroenterol Hepatol (2013) 28(Suppl 4):9–17. doi:10.1111/jgh.12294
26. Krajmalnik-Brown R, Ilhan ZE, Kang DW, DiBaise JK. Effects of gut microbes on nutrient absorption and energy regulation. Nutr Clin Pract (2012) 27(2):201–14. doi:10.1177/0884533611436116
27. Dupont A, Heinbockel L, Brandenburg K, Hornef MW. Antimicrobial peptides and the enteric mucus layer act in concert to protect the intestinal mucosa. Gut Microbes (2014) 5(6):761–5. doi:10.4161/19490976.2014.972238
28. Mukherjee S, Hooper LV. Antimicrobial defense of the intestine. Immunity (2015) 42(1):28–39. doi:10.1016/j.immuni.2014.12.028
29. Hofmann AF, Eckmann L. How bile acids confer gut mucosal protection against bacteria. Proc Natl Acad Sci U S A (2006) 103(12):4333–4. doi:10.1073/pnas.0600780103
30. Ostaff MJ, Stange EF, Wehkamp J. Antimicrobial peptides and gut microbiota in homeostasis and pathology. EMBO Mol Med (2013) 5(10):1465–83. doi:10.1002/emmm.201201773
31. Kopp ZA, Jain U, Van Limbergen J, Stadnyk AW. Do antimicrobial peptides and complement collaborate in the intestinal mucosa? Front Immunol (2015) 6:17. doi:10.3389/fimmu.2015.00017
32. Howe SE, Lickteig DJ, Plunkett KN, Ryerse JS, Konjufca V. The uptake of soluble and particulate antigens by epithelial cells in the mouse small intestine. PLoS One (2014) 9(1):e86656. doi:10.1371/journal.pone.0086656
33. McDole JR, Wheeler LW, McDonald KG, Wang B, Konjufca V, Knoop KA, et al. Goblet cells deliver luminal antigen to CD103+ dendritic cells in the small intestine. Nature (2012) 483(7389):345–9. doi:10.1038/nature10863
34. Knoop KA, McDonald KG, McCrate S, McDole JR, Newberry RD. Microbial sensing by goblet cells controls immune surveillance of luminal antigens in the colon. Mucosal Immunol (2015) 8(1):198–210. doi:10.1038/mi.2014.58
35. Birchenough GM, Johansson ME, Gustafsson JK, Bergstrom JH, Hansson GC. New developments in goblet cell mucus secretion and function. Mucosal Immunol (2015) 8(4):712–9. doi:10.1038/mi.2015.32
36. Knoop KA, McDonald KG, Kulkarni DH, Newberry RD. Antibiotics promote inflammation through the translocation of native commensal colonic bacteria. Gut (2016) 65(7):1100–9. doi:10.1136/gutjnl-2014-309059
37. Rescigno M, Urbano M, Valzasina B, Francolini M, Rotta G, Bonasio R, et al. Dendritic cells express tight junction proteins and penetrate gut epithelial monolayers to sample bacteria. Nat Immunol (2001) 2(4):361–7. doi:10.1038/86373
38. Vallon-Eberhard A, Landsman L, Yogev N, Verrier B, Jung S. Transepithelial pathogen uptake into the small intestinal lamina propria. J Immunol (2006) 176(4):2465–9. doi:10.4049/jimmunol.176.4.2465
39. Diehl GE, Longman RS, Zhang JX, Breart B, Galan C, Cuesta A, et al. Microbiota restricts trafficking of bacteria to mesenteric lymph nodes by CX(3)CR1(hi) cells. Nature (2013) 494(7435):116–20. doi:10.1038/nature11809
40. Niess JH, Brand S, Gu X, Landsman L, Jung S, McCormick BA, et al. CX3CR1-mediated dendritic cell access to the intestinal lumen and bacterial clearance. Science (2005) 307(5707):254–8. doi:10.1126/science.1102901
41. Gross M, Salame TM, Jung S. Guardians of the gut – murine intestinal macrophages and dendritic cells. Front Immunol (2015) 6:254. doi:10.3389/fimmu.2015.00254
42. Kim KW, Vallon-Eberhard A, Zigmond E, Farache J, Shezen E, Shakhar G, et al. In vivo structure/function and expression analysis of the CX3C chemokine fractalkine. Blood (2011) 118(22):e156–67. doi:10.1182/blood-2011-04-348946
43. Woof JM, Russell MW. Structure and function relationships in IgA. Mucosal Immunol (2011) 4(6):590–7. doi:10.1038/mi.2011.39
44. Brenchley JM, Douek DC. Microbial translocation across the GI tract. Annu Rev Immunol (2012) 30:149–73. doi:10.1146/annurev-immunol-020711-075001
45. Desai MS, Seekatz AM, Koropatkin NM, Kamada N, Hickey CA, Wolter M, et al. A dietary fiber-deprived gut microbiota degrades the colonic mucus barrier and enhances pathogen susceptibility. Cell (2016) 167(5):1339–53 e21. doi:10.1016/j.cell.2016.10.043
46. Johansson ME, Jakobsson HE, Holmen-Larsson J, Schutte A, Ermund A, Rodriguez-Pineiro AM, et al. Normalization of host intestinal mucus layers requires long-term microbial colonization. Cell Host Microbe (2015) 18(5):582–92. doi:10.1016/j.chom.2015.10.007
47. Petersson J, Schreiber O, Hansson GC, Gendler SJ, Velcich A, Lundberg JO, et al. Importance and regulation of the colonic mucus barrier in a mouse model of colitis. Am J Physiol Gastrointest Liver Physiol (2011) 300(2):G327–33. doi:10.1152/ajpgi.00422.2010
48. Wrzosek L, Miquel S, Noordine ML, Bouet S, Joncquel Chevalier-Curt M, Robert V, et al. Bacteroides thetaiotaomicron and Faecalibacterium prausnitzii influence the production of mucus glycans and the development of goblet cells in the colonic epithelium of a gnotobiotic model rodent. BMC Biol (2013) 11:61. doi:10.1186/1741-7007-11-61
49. Forder RE, Howarth GS, Tivey DR, Hughes RJ. Bacterial modulation of small intestinal goblet cells and mucin composition during early posthatch development of poultry. Poult Sci (2007) 86(11):2396–403. doi:10.3382/ps.2007-00222
50. Wlodarska M, Willing B, Keeney KM, Menendez A, Bergstrom KS, Gill N, et al. Antibiotic treatment alters the colonic mucus layer and predisposes the host to exacerbated Citrobacter rodentium-induced colitis. Infect Immun (2011) 79(4):1536–45. doi:10.1128/IAI.01104-10
51. Johansson ME, Gustafsson JK, Sjoberg KE, Petersson J, Holm L, Sjovall H, et al. Bacteria penetrate the inner mucus layer before inflammation in the dextran sulfate colitis model. PLoS One (2010) 5(8):e12238. doi:10.1371/journal.pone.0012238
52. Wang L, Fouts DE, Starkel P, Hartmann P, Chen P, Llorente C, et al. Intestinal REG3 lectins protect against alcoholic steatohepatitis by reducing mucosa-associated microbiota and preventing bacterial translocation. Cell Host Microbe (2016) 19(2):227–39. doi:10.1016/j.chom.2016.01.003
53. Yan AW, Fouts DE, Brandl J, Starkel P, Torralba M, Schott E, et al. Enteric dysbiosis associated with a mouse model of alcoholic liver disease. Hepatology (2011) 53(1):96–105. doi:10.1002/hep.24018
54. Hooper LV, Stappenbeck TS, Hong CV, Gordon JI. Angiogenins: a new class of microbicidal proteins involved in innate immunity. Nat Immunol (2003) 4(3):269–73. doi:10.1038/ni888
55. Eliakim R, Mahmood A, Alpers DH. Rat intestinal alkaline phosphatase secretion into lumen and serum is coordinately regulated. Biochim Biophys Acta (1991) 1091(1):1–8. doi:10.1016/0167-4889(91)90213-H
56. Nakano T, Inoue I, Alpers DH, Akiba Y, Katayama S, Shinozaki R, et al. Role of lysophosphatidylcholine in brush-border intestinal alkaline phosphatase release and restoration. Am J Physiol Gastrointest Liver Physiol (2009) 297(1):G207–14. doi:10.1152/ajpgi.90590.2008
57. Malo MS, Alam SN, Mostafa G, Zeller SJ, Johnson PV, Mohammad N, et al. Intestinal alkaline phosphatase preserves the normal homeostasis of gut microbiota. Gut (2010) 59(11):1476–84. doi:10.1136/gut.2010.211706
58. Manichanh C, Rigottier-Gois L, Bonnaud E, Gloux K, Pelletier E, Frangeul L, et al. Reduced diversity of faecal microbiota in Crohn’s disease revealed by a metagenomic approach. Gut (2006) 55(2):205–11. doi:10.1136/gut.2005.073817
59. Kaliannan K, Wang B, Li XY, Kim KJ, Kang JX. A host-microbiome interaction mediates the opposing effects of omega-6 and omega-3 fatty acids on metabolic endotoxemia. Sci Rep (2015) 5:11276. doi:10.1038/srep11276
60. Bates JM, Akerlund J, Mittge E, Guillemin K. Intestinal alkaline phosphatase detoxifies lipopolysaccharide and prevents inflammation in zebrafish in response to the gut microbiota. Cell Host Microbe (2007) 2(6):371–82. doi:10.1016/j.chom.2007.10.010
PubMed Abstract | CrossRef Full Text | Google Scholar
61. Yang Y, Millan JL, Mecsas J, Guillemin K. Intestinal alkaline phosphatase deficiency leads to lipopolysaccharide desensitization and faster weight gain. Infect Immun (2015) 83(1):247–58. doi:10.1128/IAI.02520-14
62. Bates JM, Mittge E, Kuhlman J, Baden KN, Cheesman SE, Guillemin K. Distinct signals from the microbiota promote different aspects of zebrafish gut differentiation. Dev Biol (2006) 297(2):374–86. doi:10.1016/j.ydbio.2006.05.006
63. Liu W, Hu D, Huo H, Zhang W, Adiliaghdam F, Morrison S, et al. Intestinal alkaline phosphatase regulates tight junction protein levels. J Am Coll Surg (2016) 222(6):1009–17. doi:10.1016/j.jamcollsurg.2015.12.006
64. Bevins CL, Salzman NH. Paneth cells, antimicrobial peptides and maintenance of intestinal homeostasis. Nat Rev Microbiol (2011) 9(5):356–68. doi:10.1038/nrmicro2546
65. Gallo RL, Hooper LV. Epithelial antimicrobial defence of the skin and intestine. Nat Rev Immunol (2012) 12(7):503–16. doi:10.1038/nri3228
66. Williams JM, Duckworth CA, Burkitt MD, Watson AJ, Campbell BJ, Pritchard DM. Epithelial cell shedding and barrier function: a matter of life and death at the small intestinal villus tip. Vet Pathol (2015) 52(3):445–55. doi:10.1177/0300985814559404
67. Sommer F, Nookaew I, Sommer N, Fogelstrand P, Backhed F. Site-specific programming of the host epithelial transcriptome by the gut microbiota. Genome Biol (2015) 16:62. doi:10.1186/s13059-015-0614-4
68. Reikvam DH, Erofeev A, Sandvik A, Grcic V, Jahnsen FL, Gaustad P, et al. Depletion of murine intestinal microbiota: effects on gut mucosa and epithelial gene expression. PLoS One (2011) 6(3):e17996. doi:10.1371/journal.pone.0017996
69. Stedman A, Nigro G, Sansonetti PJ. [Microbiota-intestinal stem cells dialog: a key element for intestinal regeneration]. Med Sci (Paris) (2016) 32(11):983–90. doi:10.1051/medsci/20163211014
70. Hughes KR, Harnisch LC, Alcon-Giner C, Mitra S, Wright CJ, Ketskemety J, et al. Bifidobacterium breve reduces apoptotic epithelial cell shedding in an exopolysaccharide and MyD88-dependent manner. Open Biol (2017) 7(1):160155. doi:10.1098/rsob.160155
71. Williams JM, Duckworth CA, Watson AJ, Frey MR, Miguel JC, Burkitt MD, et al. A mouse model of pathological small intestinal epithelial cell apoptosis and shedding induced by systemic administration of lipopolysaccharide. Dis Model Mech (2013) 6(6):1388–99. doi:10.1242/dmm.013284
72. Neal MD, Sodhi CP, Jia H, Dyer M, Egan CE, Yazji I, et al. Toll-like receptor 4 is expressed on intestinal stem cells and regulates their proliferation and apoptosis via the p53 up-regulated modulator of apoptosis. J Biol Chem (2012) 287(44):37296–308. doi:10.1074/jbc.M112.375881
73. Kaiko GE, Ryu SH, Koues OI, Collins PL, Solnica-Krezel L, Pearce EJ, et al. The colonic crypt protects stem cells from microbiota-derived metabolites. Cell (2016) 167(4):1137. doi:10.1016/j.cell.2016.10.034
74. Ulluwishewa D, Anderson RC, McNabb WC, Moughan PJ, Wells JM, Roy NC. Regulation of tight junction permeability by intestinal bacteria and dietary components. J Nutr (2011) 141(5):769–76. doi:10.3945/jn.110.135657
75. Donato KA, Gareau MG, Wang YJ, Sherman PM. Lactobacillus rhamnosus GG attenuates interferon-{gamma} and tumour necrosis factor-alpha-induced barrier dysfunction and pro-inflammatory signalling. Microbiology (2010) 156(Pt 11):3288–97. doi:10.1099/mic.0.040139-0
76. Johnson-Henry KC, Donato KA, Shen-Tu G, Gordanpour M, Sherman PM. Lactobacillus rhamnosus strain GG prevents enterohemorrhagic Escherichia coli O157: H7-induced changes in epithelial barrier function. Infect Immun (2008) 76(4):1340–8. doi:10.1128/IAI.00778-07
77. Miyauchi E, Morita H, Tanabe S. Lactobacillus rhamnosus alleviates intestinal barrier dysfunction in part by increasing expression of zonula occludens-1 and myosin light-chain kinase in vivo. J Dairy Sci (2009) 92(6):2400–8. doi:10.3168/jds.2008-1698
78. Seth A, Yan F, Polk DB, Rao RK. Probiotics ameliorate the hydrogen peroxide-induced epithelial barrier disruption by a PKC- and MAP kinase-dependent mechanism. Am J Physiol Gastrointest Liver Physiol (2008) 294(4):G1060–9. doi:10.1152/ajpgi.00202.2007
79. Resta-Lenert S, Barrett KE. Live probiotics protect intestinal epithelial cells from the effects of infection with enteroinvasive Escherichia coli (EIEC). Gut (2003) 52(7):988–97. doi:10.1136/gut.52.7.988
80. Ewaschuk JB, Diaz H, Meddings L, Diederichs B, Dmytrash A, Backer J, et al. Secreted bioactive factors from Bifidobacterium infantis enhance epithelial cell barrier function. Am J Physiol Gastrointest Liver Physiol (2008) 295(5):G1025–34. doi:10.1152/ajpgi.90227.2008
81. Ahrne S, Hagslatt ML. Effect of lactobacilli on paracellular permeability in the gut. Nutrients (2011) 3(1):104–17. doi:10.3390/nu3010104
82. Suzuki T. Regulation of intestinal epithelial permeability by tight junctions. Cell Mol Life Sci (2013) 70(4):631–59. doi:10.1007/s00018-012-1070-x
83. De Santis S, Cavalcanti E, Mastronardi M, Jirillo E, Chieppa M. Nutritional keys for intestinal barrier modulation. Front Immunol (2015) 6:612. doi:10.3389/fimmu.2015.00612
84. Kong J, Zhang Z, Musch MW, Ning G, Sun J, Hart J, et al. Novel role of the vitamin D receptor in maintaining the integrity of the intestinal mucosal barrier. Am J Physiol Gastrointest Liver Physiol (2008) 294(1):G208–16. doi:10.1152/ajpgi.00398.2007
85. Zhang YG, Wu S, Lu R, Zhou D, Zhou J, Carmeliet G, et al. Tight junction CLDN2 gene is a direct target of the vitamin D receptor. Sci Rep (2015) 5:10642. doi:10.1038/srep10642
86. Lam YY, Ha CW, Campbell CR, Mitchell AJ, Dinudom A, Oscarsson J, et al. Increased gut permeability and microbiota change associate with mesenteric fat inflammation and metabolic dysfunction in diet-induced obese mice. PLoS One (2012) 7(3):e34233. doi:10.1371/journal.pone.0034233
87. Chen C, Wang P, Su Q, Wang S, Wang F. Myosin light chain kinase mediates intestinal barrier disruption following burn injury. PLoS One (2012) 7(4):e34946. doi:10.1371/journal.pone.0034946
88. Earley ZM, Akhtar S, Green SJ, Naqib A, Khan O, Cannon AR, et al. Burn injury alters the intestinal microbiome and increases gut permeability and bacterial translocation. PLoS One (2015) 10(7):e0129996. doi:10.1371/journal.pone.0129996
89. Turner JR. Molecular basis of epithelial barrier regulation: from basic mechanisms to clinical application. Am J Pathol (2006) 169(6):1901–9. doi:10.2353/ajpath.2006.060681
90. Turner JR. Intestinal mucosal barrier function in health and disease. Nat Rev Immunol (2009) 9(11):799–809. doi:10.1038/nri2653
91. Cunningham KE, Turner JR. Myosin light chain kinase: pulling the strings of epithelial tight junction function. Ann N Y Acad Sci (2012) 1258:34–42. doi:10.1111/j.1749-6632.2012.06526.x
92. Kuethe JW, Armocida SM, Midura EF, Rice TC, Hildeman DA, Healy DP, et al. Fecal microbiota transplant restores mucosal integrity in a murine model of burn injury. Shock (2016) 45(6):647–52. doi:10.1097/SHK.0000000000000551
93. Chen P, Starkel P, Turner JR, Ho SB, Schnabl B. Dysbiosis-induced intestinal inflammation activates tumor necrosis factor receptor I and mediates alcoholic liver disease in mice. Hepatology (2015) 61(3):883–94. doi:10.1002/hep.27489
94. Chen P, Torralba M, Tan J, Embree M, Zengler K, Starkel P, et al. Supplementation of saturated long-chain fatty acids maintains intestinal eubiosis and reduces ethanol-induced liver injury in mice. Gastroenterology (2015) 148(1):203–14 e16. doi:10.1053/j.gastro.2014.09.014
95. Adachi Y, Moore LE, Bradford BU, Gao W, Thurman RG. Antibiotics prevent liver injury in rats following long-term exposure to ethanol. Gastroenterology (1995) 108(1):218–24. doi:10.1016/0016-5085(95)90027-6
96. Casafont Morencos F, de las Heras Castano G, Martin Ramos L, Lopez Arias MJ, Ledesma F, Pons Romero F. Small bowel bacterial overgrowth in patients with alcoholic cirrhosis. Dig Dis Sci (1996) 41(3):552–6. doi:10.1007/BF02282340
97. Mutlu EA, Gillevet PM, Rangwala H, Sikaroodi M, Naqvi A, Engen PA, et al. Colonic microbiome is altered in alcoholism. Am J Physiol Gastrointest Liver Physiol (2012) 302(9):G966–78. doi:10.1152/ajpgi.00380.2011
98. Forsyth CB, Farhadi A, Jakate SM, Tang Y, Shaikh M, Keshavarzian A. Lactobacillus GG treatment ameliorates alcohol-induced intestinal oxidative stress, gut leakiness, and liver injury in a rat model of alcoholic steatohepatitis. Alcohol (2009) 43(2):163–72. doi:10.1016/j.alcohol.2008.12.009
99. Wang Y, Kirpich I, Liu Y, Ma Z, Barve S, McClain CJ, et al. Lactobacillus rhamnosus GG treatment potentiates intestinal hypoxia-inducible factor, promotes intestinal integrity and ameliorates alcohol-induced liver injury. Am J Pathol (2011) 179(6):2866–75. doi:10.1016/j.ajpath.2011.08.039
100. Parlesak A, Schafer C, Schutz T, Bode JC, Bode C. Increased intestinal permeability to macromolecules and endotoxemia in patients with chronic alcohol abuse in different stages of alcohol-induced liver disease. J Hepatol (2000) 32(5):742–7. doi:10.1016/S0168-8278(00)80242-1
101. Uesugi T, Froh M, Arteel GE, Bradford BU, Thurman RG. Toll-like receptor 4 is involved in the mechanism of early alcohol-induced liver injury in mice. Hepatology (2001) 34(1):101–8. doi:10.1053/jhep.2001.25350
102. Elshaghabee FM, Bockelmann W, Meske D, de Vrese M, Walte HG, Schrezenmeir J, et al. Ethanol production by selected intestinal microorganisms and lactic acid bacteria growing under different nutritional conditions. Front Microbiol (2016) 7:47. doi:10.3389/fmicb.2016.00047
103. Tyler AD, Knox N, Kabakchiev B, Milgrom R, Kirsch R, Cohen Z, et al. Characterization of the gut-associated microbiome in inflammatory pouch complications following ileal pouch-anal anastomosis. PLoS One (2013) 8(9):e66934. doi:10.1371/journal.pone.0066934
104. Yu QH, Yang Q. Diversity of tight junctions (TJs) between gastrointestinal epithelial cells and their function in maintaining the mucosal barrier. Cell Biol Int (2009) 33(1):78–82. doi:10.1016/j.cellbi.2008.09.007
105. Amieva MR, Vogelmann R, Covacci A, Tompkins LS, Nelson WJ, Falkow S. Disruption of the epithelial apical-junctional complex by Helicobacter pylori CagA. Science (2003) 300(5624):1430–4. doi:10.1126/science.1081919
106. Fedwick JP, Lapointe TK, Meddings JB, Sherman PM, Buret AG. Helicobacter pylori activates myosin light-chain kinase to disrupt claudin-4 and claudin-5 and increase epithelial permeability. Infect Immun (2005) 73(12):7844–52. doi:10.1128/IAI.73.12.7844-7852.2005
107. Tetz G, Tetz V. Bacteriophage infections of microbiota can lead to leaky gut in an experimental rodent model. Gut Pathog (2016) 8:33. doi:10.1186/s13099-016-0109-1
108. Kawasaki E. Type 1 diabetes and autoimmunity. Clin Pediatr Endocrinol (2014) 23(4):99–105. doi:10.1297/cpe.23.99
109. Odenwald MA, Turner JR. Intestinal permeability defects: is it time to treat? Clin Gastroenterol Hepatol (2013) 11(9):1075–83. doi:10.1016/j.cgh.2013.07.001
110. Damci T, Nuhoglu I, Devranoglu G, Osar Z, Demir M, Ilkova H. Increased intestinal permeability as a cause of fluctuating postprandial blood glucose levels in type 1 diabetic patients. Eur J Clin Invest (2003) 33(5):397–401. doi:10.1046/j.1365-2362.2003.01161.x
111. Meddings JB, Jarand J, Urbanski SJ, Hardin J, Gall DG. Increased gastrointestinal permeability is an early lesion in the spontaneously diabetic BB rat. Am J Physiol (1999) 276(4 Pt 1):G951–7.
112. Secondulfo M, Iafusco D, Carratu R, deMagistris L, Sapone A, Generoso M, et al. Ultrastructural mucosal alterations and increased intestinal permeability in non-celiac, type I diabetic patients. Dig Liver Dis (2004) 36(1):35–45. doi:10.1016/j.dld.2003.09.016
113. El Asmar R, Panigrahi P, Bamford P, Berti I, Not T, Coppa GV, et al. Host-dependent zonulin secretion causes the impairment of the small intestine barrier function after bacterial exposure. Gastroenterology (2002) 123(5):1607–15. doi:10.1053/gast.2002.36578
114. Watts T, Berti I, Sapone A, Gerarduzzi T, Not T, Zielke R, et al. Role of the intestinal tight junction modulator zonulin in the pathogenesis of type I diabetes in BB diabetic-prone rats. Proc Natl Acad Sci U S A (2005) 102(8):2916–21. doi:10.1073/pnas.0500178102
115. Costa FR, Francozo MC, de Oliveira GG, Ignacio A, Castoldi A, Zamboni DS, et al. Gut microbiota translocation to the pancreatic lymph nodes triggers NOD2 activation and contributes to T1D onset. J Exp Med (2016) 213(7):1223–39. doi:10.1084/jem.20150744
116. Tsokos GC. Systemic lupus erythematosus. N Engl J Med (2011) 365(22):2110–21. doi:10.1056/NEJMra1100359
117. Mu Q, Zhang H, Luo XM. SLE: another autoimmune disorder influenced by microbes and diet? Front Immunol (2015) 6:608. doi:10.3389/fimmu.2015.00608
118. Nockher WA, Wigand R, Schoeppe W, Scherberich JE. Elevated levels of soluble CD14 in serum of patients with systemic lupus erythematosus. Clin Exp Immunol (1994) 96(1):15–9. doi:10.1111/j.1365-2249.1994.tb06222.x
119. Lee TP, Huang JC, Liu CJ, Chen HJ, Chen YH, Tsai YT, et al. Interactions of surface-expressed TLR-4 and endosomal TLR-9 accelerate lupus progression in anti-dsDNA antibody transgenic mice. Exp Biol Med (Maywood) (2014) 239(6):715–23. doi:10.1177/1535370214525299
120. Lee TP, Tang SJ, Wu MF, Song YC, Yu CL, Sun KH. Transgenic overexpression of anti-double-stranded DNA autoantibody and activation of Toll-like receptor 4 in mice induce severe systemic lupus erythematosus syndromes. J Autoimmun (2010) 35(4):358–67. doi:10.1016/j.jaut.2010.07.007
121. Liu B, Yang Y, Dai J, Medzhitov R, Freudenberg MA, Zhang PL, et al. TLR4 up-regulation at protein or gene level is pathogenic for lupus-like autoimmune disease. J Immunol (2006) 177(10):6880–8. doi:10.4049/jimmunol.177.10.6880
122. Levine JS, Subang R, Nasr SH, Fournier S, Lajoie G, Wither J, et al. Immunization with an apoptotic cell-binding protein recapitulates the nephritis and sequential autoantibody emergence of systemic lupus erythematosus. J Immunol (2006) 177(9):6504–16. doi:10.4049/jimmunol.177.9.6504
123. Levine JS, Subang R, Setty S, Cabrera J, Laplante P, Fritzler MJ, et al. Phospholipid-binding proteins differ in their capacity to induce autoantibodies and murine systemic lupus erythematosus. Lupus (2014) 23(8):752–68. doi:10.1177/0961203314525676
124. Tolomeo T, Rico De Souza A, Roter E, Dieude M, Amireault P, Subang R, et al. T cells demonstrate a Th1-biased response to native beta2-glycoprotein I in a murine model of anti-phospholipid antibody induction. Autoimmunity (2009) 42(4):292–5. doi:10.1080/08916930902828254
125. Lartigue A, Colliou N, Calbo S, Francois A, Jacquot S, Arnoult C, et al. Critical role of TLR2 and TLR4 in autoantibody production and glomerulonephritis in lpr mutation-induced mouse lupus. J Immunol (2009) 183(10):6207–16. doi:10.4049/jimmunol.0803219
126. Summers SA, Hoi A, Steinmetz OM, O’Sullivan KM, Ooi JD, Odobasic D, et al. TLR9 and TLR4 are required for the development of autoimmunity and lupus nephritis in pristane nephropathy. J Autoimmun (2010) 35(4):291–8. doi:10.1016/j.jaut.2010.05.004
127. Liu Y, Liao J, Zhao M, Wu H, Yung S, Chan TM, et al. Increased expression of TLR2 in CD4(+) T cells from SLE patients enhances immune reactivity and promotes IL-17 expression through histone modifications. Eur J Immunol (2015) 45(9):2683–93. doi:10.1002/eji.201445219
128. Leiss H, Niederreiter B, Bandur T, Schwarzecker B, Bluml S, Steiner G, et al. Pristane-induced lupus as a model of human lupus arthritis: evolvement of autoantibodies, internal organ and joint inflammation. Lupus (2013) 22(8):778–92. doi:10.1177/0961203313492869
129. Pawar RD, Castrezana-Lopez L, Allam R, Kulkarni OP, Segerer S, Radomska E, et al. Bacterial lipopeptide triggers massive albuminuria in murine lupus nephritis by activating toll-like receptor 2 at the glomerular filtration barrier. Immunology (2009) 128(1 Suppl):e206–21. doi:10.1111/j.1365-2567.2008.02948.x
130. Urbonaviciute V, Starke C, Pirschel W, Pohle S, Frey S, Daniel C, et al. Toll-like receptor 2 is required for autoantibody production and development of renal disease in pristane-induced lupus. Arthritis Rheum (2013) 65(6):1612–23. doi:10.1002/art.37914
131. Gallo PM, Rapsinski GJ, Wilson RP, Oppong GO, Sriram U, Goulian M, et al. Amyloid-DNA composites of bacterial biofilms stimulate autoimmunity. Immunity (2015) 42(6):1171–84. doi:10.1016/j.immuni.2015.06.002
132. Wu YW, Tang W, Zuo JP. Toll-like receptors: potential targets for lupus treatment. Acta Pharmacol Sin (2015) 36(12):1395–407. doi:10.1038/aps.2015.91
133. Teichmann LL, Schenten D, Medzhitov R, Kashgarian M, Shlomchik MJ. Signals via the adaptor MyD88 in B cells and DCs make distinct and synergistic contributions to immune activation and tissue damage in lupus. Immunity (2013) 38(3):528–40. doi:10.1016/j.immuni.2012.11.017
134. Nickerson KM, Christensen SR, Shupe J, Kashgarian M, Kim D, Elkon K, et al. TLR9 regulates TLR7- and MyD88-dependent autoantibody production and disease in a murine model of lupus. J Immunol (2010) 184(4):1840–8. doi:10.4049/jimmunol.0902592
135. Karki R, Man SM, Kanneganti TD. Inflammasomes and cancer. Cancer Immunol Res (2017) 5(2):94–9. doi:10.1158/2326-6066.CIR-16-0269
136. Karki R, Man SM, Malireddi RK, Kesavardhana S, Zhu Q, Burton AR, et al. NLRC3 is an inhibitory sensor of PI3K-mTOR pathways in cancer. Nature (2016) 540:583–87. doi:10.1038/nature20597
137. Sester DP, Sagulenko V, Thygesen SJ, Cridland JA, Loi YS, Cridland SO, et al. Deficient NLRP3 and AIM2 inflammasome function in autoimmune NZB mice. J Immunol (2015) 195(3):1233–41. doi:10.4049/jimmunol.1402859
138. Lech M, Lorenz G, Kulkarni OP, Grosser MO, Stigrot N, Darisipudi MN, et al. NLRP3 and ASC suppress lupus-like autoimmunity by driving the immunosuppressive effects of TGF-beta receptor signalling. Ann Rheum Dis (2015) 74(12):2224–35. doi:10.1136/annrheumdis-2014-205496
139. Lamprecht M, Bogner S, Schippinger G, Steinbauer K, Fankhauser F, Hallstroem S, et al. Probiotic supplementation affects markers of intestinal barrier, oxidation, and inflammation in trained men; a randomized, double-blinded, placebo-controlled trial. J Int Soc Sports Nutr (2012) 9(1):45. doi:10.1186/1550-2783-9-45
140. Hsiao EY, McBride SW, Hsien S, Sharon G, Hyde ER, McCue T, et al. Microbiota modulate behavioral and physiological abnormalities associated with neurodevelopmental disorders. Cell (2013) 155(7):1451–63. doi:10.1016/j.cell.2013.11.024
141. Round JL, Mazmanian SK. Inducible Foxp3+ regulatory T-cell development by a commensal bacterium of the intestinal microbiota. Proc Natl Acad Sci U S A (2010) 107(27):12204–9. doi:10.1073/pnas.0909122107
142. Telesford KM, Yan W, Ochoa-Reparaz J, Pant A, Kircher C, Christy MA, et al. A commensal symbiotic factor derived from Bacteroides fragilis promotes human CD39(+)Foxp3(+) T cells and Treg function. Gut Microbes (2015) 6(4):234–42. doi:10.1080/19490976.2015.1056973
143. Ochoa-Reparaz J, Mielcarz DW, Wang Y, Begum-Haque S, Dasgupta S, Kasper DL, et al. A polysaccharide from the human commensal Bacteroides fragilis protects against CNS demyelinating disease. Mucosal Immunol (2010) 3(5):487–95. doi:10.1038/mi.2010.29
Купить номер с этой статьей в pdf